 |
|
対数増殖期において細胞を合成していた栄耀細胞は種々の原因でその代謝系を芽胞への合成に変換させる。芽胞形成時には各種元素、栄養素が要求され、食品は多くの成分が含有されているので芽胞形成には好適な条件である。また古くから芽胞形成時には酵素活性の特殊性が認められている。芽胞形成に及ぼす各種の元素の影響については多くの研究者により検討されてきた。その共通点は芽胞形成における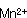 、耐熱性における 、耐熱性における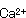 、 、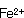 の重要性である。多くの食品にはこれらの元素は普通に含まれている。これまでの研究で芽胞が良好に形成される培地には必ずグルコースが存在している。グルコースが含まれている培地では対数増殖期のpHは急激に低下する。これはグルコースが分解されて有機酸が産生されたことによる。芽胞の形成とともに、酸素吸収が開始され、pHが上昇する。これは栄養細胞の増殖中にグルコースが完全酸化されないで、酢酸やピルビン酸のような中間代謝産物が芽胞形成とともに基質として利用され、分解されるためである。しかし食品中では種々の窒素源があるのでBacillus属の中温性菌の多くは窒素源からアンモニアが産生されてグルコースからの酸生成はこのアンモニアによりマスクされる場合が多い。 の重要性である。多くの食品にはこれらの元素は普通に含まれている。これまでの研究で芽胞が良好に形成される培地には必ずグルコースが存在している。グルコースが含まれている培地では対数増殖期のpHは急激に低下する。これはグルコースが分解されて有機酸が産生されたことによる。芽胞の形成とともに、酸素吸収が開始され、pHが上昇する。これは栄養細胞の増殖中にグルコースが完全酸化されないで、酢酸やピルビン酸のような中間代謝産物が芽胞形成とともに基質として利用され、分解されるためである。しかし食品中では種々の窒素源があるのでBacillus属の中温性菌の多くは窒素源からアンモニアが産生されてグルコースからの酸生成はこのアンモニアによりマスクされる場合が多い。
高い芽胞形成には高い窒素源が必要であるので、窒素源の多い食品はBacillus属細菌により変敗し易いと考えられる。麹等の製造においても窒素源を多くすると芽胞が多く形成することが経験的に知られている。基本的には芽胞形成は栄養細胞内に存在する酵素タンパク質が分解し、生じたアミノ酸から新たに芽胞のためのタンパク質合成が行われるのであって、外部からの補給は必要ではなく、外部からのエネルギー源または炭素源が枯渇したとき、芽胞は形成される。
食品中に多量のグルコースが存在すると芽胞の形成は停止する。これはグルコースの存在によりこれを酸化するためのを必要とする酵素系の活性が高まり、栄養細胞の増殖が高まりが多量に消費されるために芽胞の形成は行なわれない。このため糖含量の多い食品のBacillus属細菌による変敗は極めて少ない。Bacillus属細菌の芽胞は芽胞形成、発芽、発芽後成育の変化をする。芽胞形成は栄養細胞内で行なわれる現象であり、発芽及び発芽後成育は芽胞にて行なわれる。即ち栄養細胞の多い食品中では絶えず芽胞形成が行なわれ、発芽し発芽後成育を行っているのである。 |
|
 |
|
 |
|
| 食品の腐敗、変敗として起こる現象のうち、米飯、和洋菓子等の植物性加工食品や加工肉、ハムソ−セ−ジ等の動物性加工食品に粘質物の生成は普通に観察されている。Bacillus属細菌によりロ−プ生成(糸引き)とネト生成が生じる(表1)。これらの粘質物はハンバ−グ、コロッケ、パン、ケ−キ、煮豆、茹麺、カマボコ、竹輪、ナルト、ソ−セ−ジ、加工栗、おにぎり、米飯においてBacillus属細菌により生成することが古くから知られてきた。食品が Bacillus属細菌により腐敗、変敗する現象のうち、最も多いのがロ−プ生成(糸引き)とネト生成である。デ−コレ−ションケ−キ、ショ−トケ−キ、シホンケ−キ、パン、ブッセ等の洋菓子においてロ−プ現象が多くみられている。ロ−プと呼ばれるパンの変敗は、焼き上げ後12時間以内に生ずる果物の香りで始まり、これが間もなく悪臭を変わって内部が褐色未を帯び、粘ついてくる。そして、ちぎると糸を引くようになる。ロ−プ現象が生じたパンおよびその原材料から微生物を分離・同定した結果、その原因菌はB.subtilis、B.pumilus、
B.macerans、B.licheniformis、B.megateriumであった。これらの菌はいずれもロ−プが生成したデコ−レ−ションケ−キ、ショ−トケ−キ、ブッセおよびその原材料からも検出された。各種栄養培地およびスポンジ生地、ショ−トケ−キ、ブッセに上記分離菌を接種した結果、いずれの場合もロ−プ生成およびネト生成が認められた。これらの分離細菌の増殖に及ぼすpHの影響について検討した結果、pH4.0ではB.subtilis, B.stearothermophilus, B.licheniformisが増殖可能であった。またpH4.5ではB.macerans, B.polymyxaが増殖可能であった。ロ−プ生成に関与するBacillus属細菌が食品中で増殖すると増殖部位の水分が増加し、9%から14%に水分を増加した。これはB.subtilis等のロ−プ菌はでんぷんを加水分解して水分を放つためである。 ロ−プ菌の芽胞は耐熱性が高いために、パンの焙焼も生存している。しかしロ−プ現象が生じるのは菌数がある程度以上存在して一定の条件が与えられた場合のみである。酢酸やプロピオン酸塩を使用したり、パンの冷却や貯蔵中の二次汚染を防止したり、パン生地のpHを5.0〜5.5位に下げて火通りよく焼くとロ−プ現象は防止できる。ロ−プ生成はBacillus属細菌の中でも特にB.subtilis、B.licheniformisの変異株の夾膜で起こり、小麦粉のグルテンが本菌で分解され、同時にアミラ−ゼででんぷんから糖が生成してその形成を助長する。焙焼中においても生地の内部の温度が100℃を越えないために芽胞は死滅せず、洋菓子になってから適当な環境で発芽し増殖する。多くの場合まず異臭が生成し、変色が起こり、その後ロ−プや粘りが生成する。Bacillus属細菌の汚染源はほとんどが小麦粉を主とする原材料であり、小麦粉には多くのBacillus属細菌が含まれている。B.subtilis、B.pumilus、B.licheniformis
B.cereus、B.brevis、B.coagulans、B.megateriumuが検出されたという報告、B.subtilis、
B.pumilus、B.licheniformis、B.cereus、B.coagulans、B.megateriumが検出されたという報告、B.subtilis、B.licheniformis、B.cereusが多く含まれているという報告がある。冷凍生地からは
B.subtilis、B.pumilus、B.licheniformis、B.cereusが検出されている。 |
|
| 表1 Bacillus属細菌による粘質物質の生成 |
|
 |
|
ショ−トケ−キ、チョコレ−トケ−キ、バニラロ−ル、チ−ズケ−キ、チェリ−ケ−キに異臭が生成し、酸敗現象が生じた。いずれも腐敗臭はなく、酸臭が認められことからBacillus属細菌による変敗と考えられた。いずれの洋菓子においても同様な異臭と酸生成が認められたがショ−トケ−キはB.subtilis、チョコレ−トケ−キはB.cereus、バニラロ−ルはB.subtilis、チ−ズケ−キはB.subtilisとB.cereus、チェリ−ケ−キからはB.cereusが検出された。これらの洋菓子の原材料からBacillus属細菌を分離同定した結果、ショ−トケ−キ、バニラロ−ル、チ−ズケ−キから検出されたB.subtilis、チョコレ−トケ−キ、チ−ズケ−キ、チェリ−ケ−キから検出されたB.cereusは小麦粉、コ−ンスタ−チ、砂糖等に由来することが認められ、スポンジ生地焼成前に1〜2×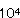 /gであり、焼成後に1〜3× /gであり、焼成後に1〜3×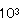 /gであった。 /gであった。
米飯はでん粉質を主成分として水分含量が多いので酸生成による異臭現象を生じやすい。20〜30℃の温度で米飯を保存すると、すえた臭いが出て、pHが低下するようになり食べられなくなるが、さらに変敗が進行すると糸を引くようになり、米粒が軟化、溶解することは経験的によく知られている。米飯の成分組成はでんぷんを主体としたものであり、100℃以上で炊きあげるため衛生的な工場で製造された米飯の変敗に関与する微生物は好気性の耐熱性芽胞菌であるBacillus属細菌がほとんどである。その原因菌としてB.megaterium, B.lentus, B.cereus, B.subtilis, B.coagulansを検出した。一般的にすえた臭気を与える原因菌としてB.subtilis, B.megaterium, B.cereus, B.mycoides, B.polymyxa, B.circulans, B.alvei, B.licheniformisが多く検出されている。
Bacillus属細菌によるフラットサワ−型変敗とは、缶の蓋底が扁平で、容器の外観に異常が認められないにもかかわらず、内容物がすっぱくなっている変敗をいう。この種の変敗は開封して初めて変敗に気づくことが多い。また容器の外観や音響検査によって変敗品の識別はできない。フラットサワ−変敗原因菌は通常工場設備の原料処理中に混入する場合が多いが、もとは砂糖、でんぷん、土壌に由来し、低酸缶詰で主に変敗が起こる。
フラットサワ−型変敗を起こしたレトルトコ−ンス−プ、レトルトコ−ンビ−フ、レトルトカレー、牛肉すき焼及び流動食の缶詰よりB.coagulansが検出され、原因菌とされた。これらの菌の芽胞は耐熱性が極めて高く、0.01Mリン酸緩衝液(pH7.0)中では、121℃におけるD値が1.4〜1.6分、Z値が7.2〜7.8であった。
プラスチックフィルムで小袋詰し、ホット充填したストレ−トス−プの混濁、異臭発生の原因した結果、変敗原因菌は原材料であるカツオパウダ−、工場落下菌より検出されたB.circulansであった。本菌は高温性であり、生育に及ぼす酸化還元電位が低く、生育最適温度は40〜50℃、生育最適pHは5〜9、100℃におけるD値は3.9〜4.0分であった。 |
|
 |
|
Bacillus属細菌により着色又は変色した食品は比較的少ない。湯殺菌包装生中華麺は、クチナシ色素又はリボフラビンを添加して着色しているが、夏期において製造後2〜3日で赤褐色斑点が生成する現象が生じた。この赤褐色斑点は保存期間の延長に伴い、徐々に大きく、濃くなる現象が生成した。この中華麺の赤褐色斑点生成現象は15℃貯蔵では5日後に、30℃貯蔵では3日後に生成し、赤褐色部の菌数は1.1×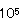 〜8.7× 〜8.7×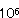 /gであり、そのほとんどがB.megateriumであった。また赤褐色部の鉄の含量は16.37〜20.61ppmであり、正常部の4.46〜6.10ppmより約4〜5倍多いことが認めた(表2)。さらに分離したB.megateriumはヒドロキザム酸の一種シゾキネンを生産していることを認めた。以上のことより湯殺菌包装生中華麺の赤褐色斑点生成現象は原材料からの一次汚染菌あるいは製造工程からの二次汚染菌であるB.megateriumがヒドロキザム酸の一種であるシゾキネンを生成し、これが微量の鉄を寄せ集めてキレ−トして赤褐色物質を生成した。鉄により清酒が着色することは、昔から良く知られていた。この原因が、大部分麹菌が生産するフェリクロ−ム類、特にフェリクリシンによることが明らかとなっている。この場合、鉄を含む着色物質であるフェリクリシンは赤褐色結晶であり、また鉄を含まないデフェリクリシンも清酒に存在している。このように清酒中には必ずデフェリクリシンが存在し、鉄の存在によりすみやかに赤褐色のフェリクロ−ムに変化することが知られている。包装生中華麺の赤褐色現象も清酒の着色と極めてよく類似していた。このため包装生中華麺の赤褐色現象を防止するためには水や食品原材料には微量の鉄が必ず含まれるためにB.megateriumを殺菌するか又はシゾキネンを生産しないようにすることが必要である。またB.megateriumはシュクロ−ス・ミネラル塩培地に添加した鉄を集積するシゾキネンを生産する。 /gであり、そのほとんどがB.megateriumであった。また赤褐色部の鉄の含量は16.37〜20.61ppmであり、正常部の4.46〜6.10ppmより約4〜5倍多いことが認めた(表2)。さらに分離したB.megateriumはヒドロキザム酸の一種シゾキネンを生産していることを認めた。以上のことより湯殺菌包装生中華麺の赤褐色斑点生成現象は原材料からの一次汚染菌あるいは製造工程からの二次汚染菌であるB.megateriumがヒドロキザム酸の一種であるシゾキネンを生成し、これが微量の鉄を寄せ集めてキレ−トして赤褐色物質を生成した。鉄により清酒が着色することは、昔から良く知られていた。この原因が、大部分麹菌が生産するフェリクロ−ム類、特にフェリクリシンによることが明らかとなっている。この場合、鉄を含む着色物質であるフェリクリシンは赤褐色結晶であり、また鉄を含まないデフェリクリシンも清酒に存在している。このように清酒中には必ずデフェリクリシンが存在し、鉄の存在によりすみやかに赤褐色のフェリクロ−ムに変化することが知られている。包装生中華麺の赤褐色現象も清酒の着色と極めてよく類似していた。このため包装生中華麺の赤褐色現象を防止するためには水や食品原材料には微量の鉄が必ず含まれるためにB.megateriumを殺菌するか又はシゾキネンを生産しないようにすることが必要である。またB.megateriumはシュクロ−ス・ミネラル塩培地に添加した鉄を集積するシゾキネンを生産する。 |
|
| 表2 包装生中華麺貯蔵中におけるBacillus属細菌による赤褐色斑点の生成と鉄含量 |
|
 |
|
古くからBacillus属細菌により離水現象が生成した食品はでん粉系食品に多い。包装五平餅が出荷後1〜2週間で完全に液化して多量の返品が出たが、その原因菌もでんぷん分解力が極めて強いB.polymyxa ,B.megaterium, B.circulans,B.mycoidesであった。Bacillus属細菌が増殖中に多量のアミラ−ゼを生産することは広く知られている。多くの研究者が食品中のでんぷん分解菌について検討し、B.subtilisが最もでんぷん分解能が大であることを報告してきた。燻製チキンロ−ルが製造後、1〜2週間で軟化し、膨張現象が生じた。これらの原因を検討した結果、軟化製品はB.macerans, B.coagulans, B.circulans, B.cereusが原因菌であり、軟化膨張品はB.licheniformis, B.brevisが原因菌であった。これらの菌はいずれもプロテア−ゼ生産能を有し、特にB.macerans, B.coagulansに高いプロテア−ゼ活性を認めた。
ようかん類には練りようかんと水ようかんがあるが、練りようかんは十分時間をかけて水分25%前後まで練り上げるもので、あん、砂糖、寒天が解け合い重量感のある旨みを持つ和菓子である。これに対して水ようかんは配合するあんを分散沸騰後、短時間で煮詰め上げるもので水分50%前後の口どけの良い清涼感のある製品である。このため練りようかんに比較して水ようかんは微生物的な変敗を受けやすい。水ようかんの変敗のほとんどは耐熱性のあるBacillus属細菌による軟化、ちじみ、膨張が中心である。軟化、ちじみ現象の生成した小倉水ようかんおよび抹茶水ようかんの湯殺菌製品(90℃、30分間)よりB.cereus, B.megaterium, B.coagulans, B.subtilis, B.polymyxaを分離するとともに軟化膨張した小倉水ようかんおよび抹茶水ようかんのレトルト殺菌製品(120℃、30分間)からB.stearothermophilusとB.circulansを分離した。
湯殺菌水ようかんでは正常製品でも離水が必ず生じており、離水率3〜5%であるが肉眼的には全く変気が認められない。しかし、離水率が20%以上となると肉眼的にも変化が認められ、この際必ずBacillus属細菌が検出されること及び無殺菌水ようかんや加熱不良水ようかんのように微生物が残存しているおそれのある場合、離水率が30%以上になることが経験的に知られていた。離水率20%以上の湯殺菌水ようかんの離水状況を(表3)に示した。
抹茶水ようかんは離水率が35〜40%と高く、次いで小倉水ようかんが30〜40%であったが、練り水ようかんは20〜35%であった。また糖濃度は正常な水ようかんはいずれもBrix42〜52%であったが離水の発生によりBrix40〜49%に低下した。また離水部分のBrixは42〜48%であった。正常な水ようかんの水分43〜48%であったが、離水により低下して36〜47%、ジェリー強度は410〜515g/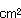 から714〜1025g/に著しく増加し、非常に硬くなった。正常な水ようかんのBacillus属菌数はいずれも300以下/gであったが、離水水ようかんでは1.1×〜3.6×/gとなった。pHは離水にかかわらず、いずれの水ようかんも製造直後とほぼ同様の6.10〜6.25の範囲であった。離水水ようかんは容器内で縮み現象を生じていた。これは水分43〜54%の水ようかんでは糖の構成比の変化に由来する浸透圧の変化に由来した(表4)。 から714〜1025g/に著しく増加し、非常に硬くなった。正常な水ようかんのBacillus属菌数はいずれも300以下/gであったが、離水水ようかんでは1.1×〜3.6×/gとなった。pHは離水にかかわらず、いずれの水ようかんも製造直後とほぼ同様の6.10〜6.25の範囲であった。離水水ようかんは容器内で縮み現象を生じていた。これは水分43〜54%の水ようかんでは糖の構成比の変化に由来する浸透圧の変化に由来した(表4)。
離水した水ようかんではショ糖が著しく減少し(98.5〜99%から7.0〜10.5%)、グルコース、フルクトースが多く検出された。離水水ようかんより9種類のBacillus属細菌を分離し同定を行った。同定した菌株はいずれもB.subtilis, B.licheniformis, B.stearothermophilusのBacillus属細菌に属した。これらの菌株を水ようかんに接種して離水率を測定した結果、いずれの菌株も20%以上の離水率を示した。特にB.subtilisを接種した水ようかんは40%の離水率を示した(表5)。20%以上の離水はBacillus属細菌の増殖により引き起こされた。すなわち同細菌が生産する酵素がショ糖を分解し、グルコースと果糖に分解されたことによりショ糖の抱水を解除したことによる。 |
|
表3 水ようかんの離水現象
表4 離水現象を生成した水ようかんの糖組成
表5 離水水ようかんより分離したBacillus属細菌接種による小倉水ようかんの離水生成 |
|
| 包装餅はポリエステル/ポリエチレン又はナイロン/ポリエチレン、ナイロン/ポリプロピレンの積層のフィルムで包装されており、かつ加熱処理されているため、従来のバラ詰餅や無包装餅に比較してその貯蔵性は大きい。一般に90〜95℃で20〜30分湯殺菌されており、100℃以上の高温度及び長時間の加熱処理では餅の「コシ」を落とし、淡黄色に着色し、さらに艶がなくなるので問題がある。しかしながら、餅製造の原料である餅玄米や精白米及び製造工程に多数の細菌、特に土壌に由来するBacillus属細菌が付着又は混入してくるため現在の加熱条件では包装餅の完全殺菌は困難であり、その保存期間は夏期特に6月〜9月に製造されたものは1〜2週間であり、極めて保存性の悪いものとなっている。9月に製造された包装餅に白色の軟化孔が生じた。この部位よりB.polymyxa, B.mycoides, B.coagulansが圧倒的に多く、B.pumilus, B.macerans, B.lentusは比較的少ないという結果及びα-アミラ−ゼ活性はB.polymyxa, B.mycoides, B.coagulansに顕著に認められたことから包装餅の軟化孔の生成原因菌はこれらの3菌株であった。包装餅の白色斑点および赤紫軟化生成原因菌はB.coagulans,B.subtilisであった。また包装生切餅の表面に微小の小孔を作り特有のムレ臭を生じる現象はB.subtilisに起因する場合もある。これらはいずれもα-アミラ−ゼ活性が極めて強く、でんぷんが分解されたことによる。この包装餅の場合、包装内が嫌気的に保持されていることより微生物の生育が阻止されている状態にある。しかし長期間保存すると菌数が増加し、軟化孔が生成する。このことは生理学的には、酸化還元電位が低い嫌気的状態とされているので、外囲の酸素分圧が高くても包装餅自体の酸化還元電位が低ければ嫌気性菌が繁殖し、逆に酸素分圧が低くても酸化還元電位が高ければBacillus属細菌も繁殖できる。細菌の繁殖と酸化還元電位の関係は多数研究されており、偏性嫌気性菌については培地の酸化還元電位が-0.1〜-0.2ボルト以下であれば、たとえ空気を吹き込んでも偏性嫌気性菌が生育する。しかし好気性菌については複雑で酸素分圧が常圧であれば、酸化還元電位が細菌の繁殖を決定するという報告や包装餅のように酸素分圧が低い状態では培地の酸素含量が細菌の繁殖を決定するという報告がされている。Bacillus属細菌は一部低電位で繁殖できる菌株と繁殖できない菌株がある。このように嫌気的な環境下での好気性細菌の繁殖については、酸素分圧と酸化還元電位の高低が複雑に影響することが考えられる。いずれにしても包装餅の貯蔵性を安全なものにするためには、まず第一に初発菌数を少なくすること、次に包装内部の酸素分圧を長期間低い状態に維持することが必要である。そのためには原料米の生菌数を及び製造工程での汚染を最小にすること、気体透過度の小さい包装材料で包装し脱気度をさらに大きくすることが必要である。 |
|
 |
|
| Bacillus属細菌の中には嫌気的に糖やでんぷんを消費し、増殖できる菌が存在する。これらの細菌は特有の発酵を行い、2,3ブタンジオール、グリセロール、炭酸ガス、乳酸、エタノール、アセトン、酢酸、ギ酸、水素を生成する。B.subtilis(グルコースを嫌気的に発酵できず、空気のあるところで2,3ブタンジオール生成), B.lichenifpormis(硝酸塩存在下で嫌気的発酵),B.polymyxa,(嫌気的に糖、でん粉を発酵して2,3ブタンジオール、エタノール、炭酸ガス、水素生産し嫌気下で窒素固定能) 、B.macerans(嫌気的に糖を発酵し、エタノール、炭酸ガス、ギ酸、アセトン、水素を生産し、嫌気下で窒素固定能)が代表的な菌である。
フルーツみつ豆において主として酵母による膨張が多く見られるが、嫌気性菌(Clostridium pastuerianum)による膨張も多く認められている。pH3.5〜3.7の包装フルーツみつ豆が膨張する現象が生じた。検討を行った結果、直接の原因菌は酪酸菌3菌株(Clostridium pastuerianum、Clostridium thermoaceticum、Clostridium butyriccum)であった。酪酸菌は発酵性の炭水化物を含まない培地中での生育は不良であるが、でん粉様の多糖を予備物質として合成する。さらに本菌の特徴は窒素を非常に活発に固定する。膨張したフルーツみつ豆より5菌株の耐熱性芽胞菌を分離した(酪酸菌3菌株、B.subtilis, B.polymyxa).フルーツみつ豆に上記5菌株を接種し、90℃で30分間殺菌後保存試験を行った結果を(表6)に示した。これらの3種類の酪酸菌単独あるいは組み合わせでは膨張は発生するが、著しい膨張とはならない。これらの酪酸菌にB.polymyxaを組み合わせた時に著しい膨張現象を認めた。包装フルーツみつ豆の製品のpHは低いためにBacillusの生育は遅いが、まずB.polymyxaが生育して酸素を消費した結果、発酵生産物を生成して嫌気状態が作り出され、その結果、酪酸菌が生育しやすくなり、ガスを生成し、膨張品を発生するとともに酸を生成してさらにpHが低下するものと考えられる。B.subtilis単独添加では上記と同様の結果を得られないので発酵性Bacillus属であるB.polumyxaのみがこのような発酵促進効果があり、膨張現象を促進する。 |
|
| 表6 包装フルーツみつ豆の膨張 |
|
 |
|
 |
|
しょうゆは発酵、熟成後に80℃で加熱処理が行われ製品化されるために、しょうゆ中の大部分の微生物は殺菌されるが、Bacillus属細菌の芽胞はこの温度では死滅せずに残存し、そのまま製品しょうゆに移行する。またBacillus属細菌が麹中に極度に増殖すると納豆麹となり醤油の品質を著しく低下させ、醤油麹中で形成された芽胞はそのまま諸味に移行して製品に移行することが確認されている。これらの細菌芽胞は高濃度の食塩を含むしょうゆ中では増殖しないので問題とはならないが、加工食品にしょうゆを使用した場合には変敗の原因菌となる場合がある。近年、製品しょうゆ中のBacillus属細菌が、加工食品中で増殖して種々の問題が生じてきた。しょうゆ製造工程で分離されたBacillus属細菌の同定では出麹、及び熟成諸味中の芽胞細菌はB.subtilisで100%であり、その理由はこの菌が蒸煮大豆中で非常に良好に生育すること及び麹菌菌叢表面上で他菌種よりも増殖率が高い、つまり麹菌による増殖抑制を受けにくいことによる。この報告では本菌の低減方法として麹菌菌糸を十分に生育させた段階で手入れを十分に行えば良く、また出麹時に存在した芽胞のうち、諸味工程を経た後でも残存する芽胞の数もB.subtilisが圧倒的に多いところから製品にも多いと考えられるとしている。出麹時の芽胞は仕込み後も芽胞として残存するが、出麹時の芽胞数7.0×/gが仕込8カ月経過後には1.0×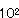 /gにまで低下した。また菌叢は出麹と同様にいずれもB.subtilisがほとんどであった。しかしこの諸味を圧搾して生醤油とし、規格調整後火入れを行い、急冷後発生したおり中の芽胞菌を分析するとB.subtilis, B.megaterium., B.circulans, B.coagulans, B.pumilusが検出された。 このことはしょうゆ工場においてはBacillus属細菌の二次汚染が多いことを示している。しょうゆ10%含有培地におけるBacillus属細菌芽胞(B.subtilis, B.cereus, B.coagulans, B.pumilus, B.circulans, B.megaterium)の増殖許容範囲は、pH4.5以上、食塩濃度12.5%以下、エタノ−ル濃度3.5%以下であった。しょうゆ中でのBacillus属細菌の形態は成分のいかんを問わず、増殖や死滅はなく、芽胞形状で生存していることが認められた。B.subtilisはしょうゆ醸造上の必要微生物といわれているが、必要以上の増殖は品質を劣化させる。このためしようゆ中の残存芽胞を最小限少なくするために、原料の完全殺菌、無菌こうじの使用、製きくにおける抑制、仕込み塩水の殺菌、除菌、仕込み容器の殺菌、工場の衛生管理が必要となる。 /gにまで低下した。また菌叢は出麹と同様にいずれもB.subtilisがほとんどであった。しかしこの諸味を圧搾して生醤油とし、規格調整後火入れを行い、急冷後発生したおり中の芽胞菌を分析するとB.subtilis, B.megaterium., B.circulans, B.coagulans, B.pumilusが検出された。 このことはしょうゆ工場においてはBacillus属細菌の二次汚染が多いことを示している。しょうゆ10%含有培地におけるBacillus属細菌芽胞(B.subtilis, B.cereus, B.coagulans, B.pumilus, B.circulans, B.megaterium)の増殖許容範囲は、pH4.5以上、食塩濃度12.5%以下、エタノ−ル濃度3.5%以下であった。しょうゆ中でのBacillus属細菌の形態は成分のいかんを問わず、増殖や死滅はなく、芽胞形状で生存していることが認められた。B.subtilisはしょうゆ醸造上の必要微生物といわれているが、必要以上の増殖は品質を劣化させる。このためしようゆ中の残存芽胞を最小限少なくするために、原料の完全殺菌、無菌こうじの使用、製きくにおける抑制、仕込み塩水の殺菌、除菌、仕込み容器の殺菌、工場の衛生管理が必要となる。
Bacillus属細菌の大半は工場の床や側溝に住み着いているので洗浄効果のあるオゾン水で芽胞ごと洗浄除去することは有効である。本菌に対しては殺菌には高濃度のオゾン水(5〜10ppm)が必要であるが洗浄には低濃度(0.3〜0.5ppm)で効果あることが多くの食品工場で確かめられている。また本菌はオゾンガス殺菌によっても減少させることが可能である。 |
|
 |
|
牛乳の生乳中に多く検出され、汚染菌量が多いのはBacillus属細菌である。生乳中のBacillusの分布を調査した結果、最も多い菌種がB.licheniformisで43.3%、次いでB.cereusの37.4%であり、その他は10%未満であったという報告や80℃で10分間加熱した生乳由来のBacillus属細菌の分布はB.subtilis30%、B.cereus19%、B.pumilus11%、B.circulans8%、B.lentus8%、B.badius3%であったという報告がある。このように生乳を汚染する好気性細菌芽胞の中ではB.subtilisとB.licheniformisが多く、耐熱性が高いのでこれらの菌を生乳の殺菌条件設定の指標菌とされる。しかし乳牛の飼料は季節により変動し、サイレ−ジ中の菌の汚染状況により指標菌は変動する。
生乳中の芽胞数の測定は採取した試料は試験管(直径18mm、長さ180mm)に採取して、80℃の恒温水槽に浸せきし、内容物が外囲温度に到達してから10分間保持して加熱処理後、標準寒天培地を用いて希釈平板法により30℃と55℃48時間培養後の集落数を計測する。
30℃で得られた芽胞数を中温性芽胞菌数、55で得られた芽胞数を高温性芽胞菌数とする。 |
|
 |
|
豆乳の原料大豆に付着耐熱性芽胞菌はほとんどBacillus属細菌であり、B.subtilis, B.coagulans, B.polymyxaが分離されており、これらの芽胞の豆乳中で加熱した時のD値は100℃ではそれぞれ18分、13.6分、及び3.1分であった。また豆腐の製造において問題となるBacillus属細菌にB.cereusがあり、市販豆乳の15%から本菌が検出され、及び豆腐とその加工品由来105株中の19株の芽胞は、105℃、10分間では殺菌できないことが報告されている。豆乳にBacillus属細菌が増殖した場合は豆乳が凝固し、B.licheniformis及びB.subtilisが増殖した場合は豆乳の表面に、それぞれ桃色及び黄褐色の菌膜を形成する。pH5.5〜7.0の豆乳中にはB.subtilis,B.licheniformis, B.circulans, B.coagulans, B.stearothermophilus等のほとんどのBacillus属細菌が発育することができる。
大豆加工品工場のBacillus属細菌の汚染は原材料に由来する工場環境からの二次汚染が中心であるのでオゾンを利用して菌数を減少させることが可能である。工場をオゾンガス(夜間)及びオゾン水(作業中)を併用すると効果がある。芽胞には低濃度オゾンでは殺菌効果が弱いが増殖が著しく抑制されるので夏季においては効果的である。
pH6.3、Brix 60%の木綿豆腐には液化品及び正常品のいずれにおいても3.5×/gのBacillus属細菌が検出された。これらはいずれも原料由来の菌であるので、原料をオゾン処理する技術も開発されている。 |
|
 |
|
煮豆は金時豆、うずら豆、いんげん豆、とら豆、大豆等の原料を用いて、砂糖、ブドウ糖、水飴、食塩、ソルビン酸等の調味液で煮込んだり、濃厚な糖液に蒸煮した豆を漬けたりして製造するものであり、ほとんどの製品が水分含量を30〜40%としたものであるために保存性が悪い。通常煮豆の保存性は1〜3日であるが、煮豆の貯蔵性を向上させる目的で、現在、ソルビン酸等の保存料を添加してプラスチックフィルムを用いた脱気包装が行われ、80〜85℃で数十分間加熱されている。それにもかかわらず煮豆製品よりB.pumilus, B.megaterium., B.cereus, B.subtilis, B.licheniformis, B.circulans, B.coagulansが検出され、腐敗した真空包装煮豆から原因菌としてB.cereusが検出された。ソルビン酸添加煮豆製品の変敗したものについてソルビン酸含量を測定すると、ソルビン酸含量が著しく低下しているものがあり、分離された菌のソルビン酸分解能を測定するとBacillus属細菌に著しいソルビン酸分解能が認められた。煮豆のネトを伴う変敗はほとんどBacillus属細菌であり、B.subtilisが主原因菌となる。一般に製造直後煮豆の菌数は〜/gであり、通常原料中にはB.circulans, B.pumilus, B.megaterium, B.cereus, B.subtilis, B.licheniformis, B.coagulans, B.stearothermophilusのBacillus属細菌が多く検出される。これらの菌はいずれも原料由来が中心であるので原料をオゾン殺菌すると効果がある。豆類の微生物はオゾン処理直後にはあまり減少しないが、貯蔵中に減少することが知られている。
オゾンは日本では食品製造用材として古くから既存食品添加物リストに挙げられ世界に先駆けて認められている。2000年8月には、米国食品医薬品局(FDA)において、食品の貯蔵及び製造に殺菌剤としてオゾンガス及びオゾン水を用いることが許可され、2001年6月に公布された。なおオゾンは食品添加物であるが、残留しないので表示の義務はない。 |
|
 |
|
ストレ−トス−プ原材料に用いるかつおパウダ−の開封品からは3.0×/gから2.7×/gのB.circulans, 未開封品からは1.0×/gから2.4×/gのB.circulansが検出された。しかし、その他の原材料である砂糖、グルタミン酸ナトリウム、食塩、味醂からはBacillus属細菌は検出されなかった。原材料を混合、撹拌して85℃まで蒸気加熱してスチ−ムをとめ90℃達温後、20分間加熱してサブタンクに移す。サブタンクは90℃に保温してあり、次にプレ−ト式熱交換機(120〜125℃)を通し、90℃に保温されたレシ−ブタンクに移行する。次の工程の充填・包装はクリ−ンル−ムの中で行い、充填温度は80〜82℃である。B.circulansは原材料の混合・撹拌工程ですでに5.0×/g存在し、サブタンクではさらに増殖して1.5×/gとなった。しかしプレ−ト式熱交換機を用いた殺菌工程で完全に死滅したが、レシ−ブタンクでは二次汚染を受けて6.0×10/gとなった。そこで工場がB.circulansに汚染されていることが考えられたので製造工程別に落下菌を測定した結果、ほとんどの工程でB.circulansを検出した。
久寿米らは100℃以下の低温加熱処理後の希釈醤油や調味液中における醤油由来細菌芽胞の挙動について検討し、希釈醤油などの成分の目安として食塩濃度7.2%以下でpH5.0以上の場合、あるいは水分活性0.92以上の場合、細菌芽胞の発芽及び生育を抑制するにはかなりの厳しい加熱条件が必要であることを報告した。また食塩濃度4%、水分活性0.95、pH5.0の成分のストレートつゆは芽胞の殺菌を目的とすることを示し、少なくとも90℃以上、場合によっては100℃以上の加熱が必要であることを示した。さらに殺菌を効果的に行うためにストレートつゆにクエン酸ナトリウムを0.2〜0.5%添加することを提案した。
|
|
 |
|
食肉は水分が50〜60%と多く、pHが5.8〜7.5と中性範囲であるので微生物は増殖しやすい。食肉は極めて多くの微生物が混入しているが、そのうちカビは表面にのみ生育し、酵母の汚染も部分的であるが、細菌は表面のみならず、内部にも侵入して増殖する。20〜30℃ではBacillus属細菌が増殖して〜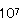 /になるとネト等が発生する。最も多く検出されるのがB.subtilisである。ソ−セ−ジ用原材料は、主原料としては豚肉が使われ、副原料としてはでんぷん、大豆たんぱく質が使用されるのでBacillus属細菌は多い。ハム製品にもBacillus属細菌が最も多い。 /になるとネト等が発生する。最も多く検出されるのがB.subtilisである。ソ−セ−ジ用原材料は、主原料としては豚肉が使われ、副原料としてはでんぷん、大豆たんぱく質が使用されるのでBacillus属細菌は多い。ハム製品にもBacillus属細菌が最も多い。
燻製チキンロ−ルは豚肉を原料として製造されたベ−コンを芯にしてチキンをロ−ル状に巻いた後、調味液に2℃で12時間漬込み、取り出した後つるし、蒸気で最終78℃になるように加熱し調理を行う。次にスモ−クハウスに入れ、45〜60℃で3時間燻煙処理を行った後、真空パックを行っている。真空パック後は100℃で3分間殺菌処理を行ってから冷凍し貯蔵保存している。そして出荷時に常温にもどし2〜3日以内に冷蔵される。なお豚ベ−コンの製造方法は豚バラ肉を12時間タンブリング後、1週間冷蔵してねかす。その後スモ−クハウスに入れ、12時間燻煙処理を行い、この間に中心部温度が80℃に到達する。燻煙処理後、スライスし冷凍されている。製造工程におけるBacillus属細菌の変化について検討した結果、いずれの工程においても7.5×10/gから3.6×/g検出された。豚ベ−コンを芯にしてチオキンロ−ル状に巻く工程で7.5×10/gであり、調味液浸漬工程で1.5×/gとなった。調味液浸漬温度は2℃であるが、12時間行うために増殖したと考えられた。さらに調味液浸漬終了後、つるして液を切り、最終78℃程度になるようにスチ−ムで加熱調理する工程で2.0×/gとなり、45〜60℃で3時間行う燻煙処理により5.0×/gとなった。燻煙処理後、冷却を行い、真空パックを行うが真空パック工程で二次汚染を受けて3.5×/gとなり、次の加熱殺菌工程(100℃、3分)により2.7×/gとなり、冷凍と解凍を繰り返すうちに増加し、出荷時に
3.6×/g となり、通常の正常製品からはB.megateriumが多く検出された。
食肉缶詰の殺菌は、他の缶詰と異なり加熱により品質が劣化するため、比較的緩和されている。このような環境で商業的無菌状態を確保しているのは、亜硝酸ナトリウム及び食塩等の静菌作用によるものと考えられ、Perigo効果として知られている。このためClostridium属細菌による変敗はほとんどないが、B.licheniformisによる変敗が多く見いだされている。その理由として、一般にB.licheniformisは、原料肉に付着する芽胞細菌の中で最も多く含まれている菌で、殺菌後に生残する可能性が高いこと、合成培地中において3〜5%濃度食塩の存在が耐熱性を増大させること及び亜硝酸ナトリウム単独では500ppmまで添加してもほとんど耐熱性に影響を与えないことによる。
現在、日本を初めとする多くの国で工場殺菌や食品原材料の殺菌にオゾンが利用されている。
|
|
|